Annual Report of the Bean Improvement Cooperative (1998) 41: 105-106.
Identification of polytene chromosomes of Phaseolus coccineus on
the basis of centromeric heterochromatin morphology
Nenno,
M., Nagl,
W.
Division
of Cell Biology, The University of Kaiserslautern, P.O. Box 30 49,
D-67653 Kaiserslautern, Germany
Introduction
Since the discovery of polytene chromosomes in the embryo suspensor of
Phaseolus coccineus (Nagl 1962a) attempts were made to accurately
describe each chromosome (Nagl 1962b, 1965). A preliminary idiogram based
on total chromosome length and the distribution of euchromatic and heterochromatic
segments was first presented by Nagl (1967). Later, Nagl (1974) pointed
out, however, that the length can vary considerably depending on the level
of endopolyploidy, the state of condensation and the degree of stretching
during preparation. Therefore it was necessary to improve the description
of the karyotype and to find a more reliable and constant feature in order
to identify each chromosome of a given nucleus. We report here the first
results of the identification of the polytene chromosomes of P. coccineus
based on the analysis of centromeric heterochromatin morphology after pepsin
pretreatment and DAPI staining.
Material and methods
Seeds of Phaseolus coccineus cv. Preisgewinner
were grown in the Botanical Garden of the University of Kaiserslautern.
Embryo suspensors were taken from seeds of unripe pods and preparations
of polytene chromosomes were made from the biggest basal suspensor cells
(Nagl 1996). After enzymatic maceration the nuclei were gently squeezed
out from single cells to get rid of the cell wall. Additionally, the extracted
nuclei were incubated in 45% acetic acid before squashing in order to dissolve
the cytoplasm to some extent, and thus improving the spreading of chromosomes.
After removing the cover slip and air drying, the preparations were pretreated
in the same way as was carried out for fluorescence in situ hybridization
(Nenno et al. 1994). Chromosomes were then stained with DAPI. Pictures
from chromosomes of ten well preserved nuclei were taken by a cooled CCD
camera at high resolution and analyzed by using an image analysis system
(IPLab Spectrum).
Results and Discussion
The analysis of polytene chromosomes from the embryo suspensor of P.
coccineus after pepsin pretreatment and DAPI banding made it possible
to give a better description of individual chromosomes (Fig.1).
The new key feature is the centromeric heterochromatin morphology (CHM).
Two characteristics make the CHM useful for chromosome identification.
Firstly, it is less sensible to variations in level of endopolyploidy and
the degree of stretching during preparation. Secondly, the CHM of each
of the 11 chromosome pairs can be individually identified by its pattern
of at least one bright DAPI band as well as by the position of the centromere
and the presence of secondary constrictions.
For the construction of an idiogram, traditionally, the total length
and the ratio of the short and the long arms are relevant. In the polytene
chromosomes of P. coccineus these parameters, however, vary significantly
due to the state of condensation and the degree of stretching during preparation.
Especially, the euchromatin behaves like a elastic band. For this reason,
the arrangement of chromosomes labeled C-J in Fig. 1
is arbitrary, and the euchromatic parts are marked with dashed lines. Theses
chromosomes are metacentric to submetacentric. Chromosome A is the longest
and is telocentric. B is the next longest chromosome and the only one which
is generally submetacentric. Chromosome K is the smallest of the complement
and is acrocentric. Since the telomeres mostly appear as clusters of heterochromatin
knobs they are symbolized by triple dots in Fig. 1.
Besides the new characterization of the chromosomes by the CHM there
are two striking differences between the new idiogram and that previously
proposed (Nagl 1967). The new idiogram shows three chromosome carrying
a NOR (Fig. 1 A, C, K). This was also demonstrated by silver-staining (Schweizer
and Ambros 1979) and by fluorescence in situ hybridization (Nenno
et al. 1994). Furthermore, the existence of the formerly named 'SAT-chromosomes'
1 (S1) and 5 (S2) (Nagl 1965, 1967) could not be
confirmed. Our present results indicate, that chromosome fragments sticking
to the nucleolar material were obviously misinterpreted as 'chromosome
satellites'. These fragments seem to arise from the small centromeric heterochromatin
block close to the NOR on the short arm of chromosome K, from which the
long arm readily detaches (Fig.1).
The problem, however, that not all of the eleven pairs can be recognized
equally well is not yet solved. Although, eight pairs of homologous can
now be recognized quite easily, the identification of chromosome H, I,
and J is more difficult. Nevertheless, the preliminary results presented
here are encouraging and suggest that in a near future it will be possible
to produce a new, high resolution karytotype for Phaseolus coccineus.
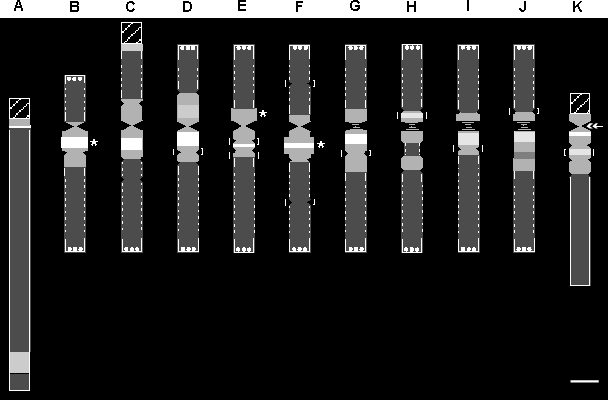
Figure 1: Idiogram of the polytene chromosomes of Phaseolus
coccineus on the basis of centromeric heterochromatin morphology (CHM).
Hatched boxes:NORs; Triple dots: Telomeric heterochromatin:
Horizontal hatching: Centromeres which are not always visible; Dark
: Euchromatin; Centromeric Dark gray: heterochromatin (CH); Gray
or white: DAPI-bands within the CH, according to their relative
staining intensity. Square brackets: structures which are not always
present; Double headed arrow: the position in K, where the long
arm detaches readily; Asterisks: regions which normally are wider
than the rest of the chromosome. Bar represents 10 µm.
The idiogram and a complete karyotype for the polytene chromosomes is
available in BeanRef
(Nenno and Nagl 1996) on the WWW at the URL:
http://www.ba.cnr.it/Beanref [
see section: Cytogenetics]
Acknowledgment
This work was supported by the Deutsche Forschungsgemeinschaft (DFG, grant
no. Na 107/10-2).
References
Nagl W. (1962a) Naturwiss. 49: 261-262.
Nagl W. (1962b) Österr. Bot. Z. 109: 432-494.
Nagl W. (1965) Chromosoma 16: 511-520.
Nagl W. (1974) Z. Pflanzenphysiol. 73: 1-44.
Nagl W. (1996) In: Fukui K. and Nakayama S. (Eds.) Boca Raton, CRC
Press pp. 51-83.
Nenno M., Schumann K., Nagl W. (1994) Genome 37:1018-1021.
Nenno M. and Nagl W. (1996) Annu. Rep. Bean Improv. Coop. 39:
200.
Schweizer D. and Ambros P. (1979) Pl. Syst. Evol. 132:27-51.