From a structural point of view polytene chromosomes can be defined as a special case of chromosomes characterized by (i) a high content of DNA and (ii) particular chromatin structures related to their high DNA contents (see below).
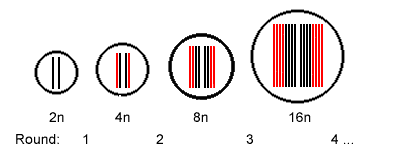
As a consequence of the enlarged amount of DNA polytene chromosomes are considerably longer than chromosomes from cells with regular DNA content (chromosomes in metaphase). Because of their larger size compared to metaphase chromosomes polytene chromosomes were called in older literature as giant chromosomes.
Polytene chromosomes can be found in many different species. From simple organisms to plants and animals. Find an extensive description, list of species, and references in Zhimulev (1996) and Sorsa (1988)
A common structural feature of all polytene chromosomes is that
their many chromatin filaments (chromatids) are laying -more or less
parallel- side by side.
The reason for the higher number of chromtids
is a particular mechanism of replication of DNA content where after
each round of duplication the resulting new chromatids are not
separated and but remain within the same cell (endoreduplication).
The result is a cell that is doubling its DNA content with each
replication round. The number of replications rounds can be as high
as 10 and higher resulting 1024, 4096 or even 16384 times the
regular DNA content.
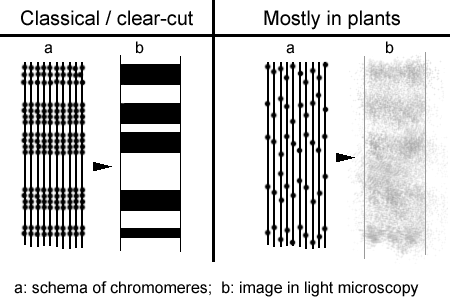
The second structural feature is a kind of transversal banding pattern (similar to a bar code) along the chromosome that can be easily observed in light microscopy. Comparing this banding pattern of lighter and darker chromosomal segments both in light and electron microscopy, it could be shown that the darker segments are represented by chromatin that is packaged relatively dense, while the brighter segments (interbands) are made up by chromatin that is packed much more loosely.
Polytene chromosomes displaying a very clear-cut banding pattern are called classical polytene chromosomes. The most famous classical polytene chromosomes can befound in Diptera (e.g. the fruit fly Drosophila melanogaster, Chironomus tentans, Anopheles) and in Ciliates (e.g. Stylonychia, Euplotes, Oxytricha).
Polytene chromosomes of plants do rarely display a clear banding pattern. A possible explanation is that chromomeres in this case are not lined up exactly in parallel and as a consequence the more dense chromatin parts are not lying regular enough side-by-side to form a clear-cut banding pattern (see figure below).
Polytene chromosomes are chromatin in interphase and as such showing transcriptional activity. If a gene is transcribed in very high quantity the surrounding chromatin is loosing its dense packaging and the region around this gene becomes larger. This can be observed in light microscopy as a local widening of a short chromosomal segment and this structure is called a puff.
Some puffs (Balbiani rings) are always found at the same chromosomal position and named after Balbiani who discovered them for the first time. The majority of puffs, however, are visible only at certain times of the development of an organism and they appear at different positions on the polytene chromosomes. This is explained by the fact that at different times during the development different genes are active. Furthermore, the activity of genes can be different from one tissue to another one.
The first description of polytene chromosomes in plants originate, like those in Diptera, from the end of the ninetheenth centry. Strasburger (1887) discovered them in the embryo sack of Fritillaria imperialis and Osterwalder (1898) in the antipods of Aconitum. Osterwalder was the also the first who recognized the similarity between these chromosomes found in plants and those chromosomes discovered just a few years earlier in the salivary glands of the diptera Chironomus by Balbiani (1881). During the following decades, however, the finding of polytene chromosomes in plants got lost and were re-discovered by the Tschmermak-Woess and Hasitschka. They found them e.g. in the integument epidermis of Melandrium viscosum, the trichomes ... of Bryonia, Papaver rhoeas and the antipods of Aconitum, which were studied already before by Osterwalder (Tschermak-Woess 1956, Hasitschka 1956).
The term polytene chromosomes, however, was used only in quotation marks for these chromosomes in plants since they did not not always display the trasversal banding patterns and the length ratio between mitotic to polytene chromosomes of about 1:10 was rather low compared to those of Diptera. More common was the name giant chromosomes
In the genus Phaseolus large polytene chromosomes with transversal banding pattern were found in the large cells of the embryo suspensor of P. coccineus and P. vulgaris (Nagl 1962a; Nagl 1969b). Although the tendency for polyteny was found also in six other species of Phaseolus the one of P. coccineus and P. vulgaris show the highest degree of polyteny and their chromosomes are most compact from ten species studied (Nagl, 1974).
In the tapetum of anthers of P. coccineus and Vigna unguiculata have been found smaller and less compact polytene chromosomes (Guerra und Carvalheira 1994). A recent review describes the history and current state of fluorescence in situ hybridizions in Phaseolus and Vigna (Guerra 2001).
Ammermann D. (1971) Morphology and development of the macronuclei of the ciliates Stylonychia mytilus and Euplotes aediculatus. Chromosoma 33: 209-238. Contains fotos and gives explanation of the temporary occurrence of polytene chromosomes in two cilitae species.
Balbiani E. G. (1881) [French] Sur la structure du noyau des cellules salivaires chez les larves de Chironomus. Zool. Anz. 4: 637-641, 662-666. First report on polytene chromosomes in Diptera
Carvalheira, G. M. G. (2000) Plant polytene chromosomes [Review]Genetics and Molecular Biology: 23(4):1043-1050.
Guerra M. (2001) Fluorescent in situ hybridization in plant polytene chromosomes. Methods Cell Sci.: 23(1-3):133-138.
Guerra M., Carvalheira G. M. G. (1994) Occurrence of polytene chromosomes in the anther tapetum of Vigna unguiculata (L) Walp. J. Hered. 85: 43-46.
Hasitschka G. (1956) Bildung von Chromosomenbündeln nach Art der Speicheldrüsenchromosomen, spiralisierte Ruhekernchromosomen und andere Struktureigentümlichkeiten in den endopolyploiden Riesenkernen der Antipoden von Papaver rhoeas. Chromosoma 8: 87-113.
Nagl W. (1962) [German] 4096-Ploidie und 'Riesenchromosomen' im Suspensor von Phaseolus coccineus. Naturwissenschaften 49: 261-262. First report on occurrence of Polytene chromosomes in the genus Phaseolus
Nagl W. (1969) Banded polytene chromosomes in the legume Phaseolus vulgaris. Nature 221: 70-71.
Nagl W. (1974) The Phaseolus suspensor and its polytene chromosomes. Z. Pflanzenphysiol. 73: 1-44. First review of Phaseolus polytene chromosomes.
Nagl W. (1981) Polytene Chromosomes of Plants. Int. Rev. Cytol. 73: 21-53. Updated review of Phaseolus polytene chromosomes.
Osterwalder A. (1898) [German] Beiträge zur Embryologie von Aconitum napellus L. Flora 85: 254.
Sorsa V. (1988) Polytene Chromosomes in Genetic Research. Chichester, Ellis Harwood Limited.
Strasburger E. (1887) [German] Das Botanische Practicum - Anleitung zum Selbststudium der mikroskopischen Botanik. 2. Umgearbeitete Auflage. Jena, Gustav Fischer. First documentation of polytene chromosomes in plants (Fritillaria imperialis). However, it is a textbook written in German and probably for this reasons it didn't attract a lot of attention.
Tschermak-Woess E. (1956) Notizen über die Riesenkerne und 'Riesenchromosomen' in den Antipoden von Aconitum. Chromosoma 8: 114-134.
Zhimulev I. F. (1996) Morphology and structure of polytene chromosomes. In: Hall J. C., Dunlap J. C. Advances in Genetics, Vol 34. San Diego, Academic Press Inc. 490pp. A detailed Monography about polytene chromosomes containing fotos, schemas and species list together with many references.